













모두가 양돈 현장에서 PRRS를 여러 차례 겪어 봤지만, 아직도 PRRS 컨트롤에 대한 '정답'은 없습니다. 오랜 기간 양돈농가를 괴롭혀온 만큼 PRRS에 대한 오해와 편견이 많이 쌓여있는 현실입니다. 'PRRS의 모든 지식'(총 15화)을 통해 우리 농장에 맞는 PRRS 컨트롤의 '해답'을 발견할 수 있길 기대합니다. 본 기고글은 HIPRA 본사에서 출간한 'The book for PRRS Knowledge"' 내용을 번역·정리한 것입니다.
PRRS 바이러스는 어떻게 분류될까?
최근 바이러스 분류 국제위원회(ICTV)는 PRRS 바이러스의 두 유전형(genotype)을 ‘PRRSV1’과 ‘PRRSV2’라는 두 개의 서로 다른 종(species)으로 재분류했습니다. PRRSV1과 PRRSV2는 모두 Poraterivirus 속(genus)에 포함됩니다. Poraterivirus는 다른 네 가지 속(genus)의 바이러스들과 함께 Arteriviridae 과(family)에 포함되며, Arteriviridae는 Mesoniviridae, Roniviridae, Coronaviridae 등의 다른 과(family)의 바이러스들과 함께 Nidovirales 목(order)을 구성합니다.

Arteriviridae 과(family)에 속하는 바이러스들 사이의 유전적 상동성은 대략 40-70%로 알려져 있습니다. 이들은 같은 목(order)의 바이러스들과 진화 과정상 관계성이 있긴 하지만, 아래와 같은 몇 가지 고유한 특성을 나타냅니다.
● 고유한 유전자 구성과 크기(12.7~15.7kb)
● 유전적 변이가 활발하게 나타남
● 형태학적으로 고유한 모양을 가진 바이러스 입자
● 세포 친화성: 주머니쥐에서의 신경병 바이러스(WPDV, Wobbly possum disease virus) 및 유인원에 감염되는 몇 종류의 바이러스를 제외하고, 모두 폐포 대식세포(alveolar macrophage)에서 증식하는 특징을 갖고 있음
● 지속감염 및 무증상 감염을 일으킬 수 있음
PRRS 바이러스는 어떻게 구성되어 있을까?
PRRS 바이러스의 유전체 구성
PRRS 바이러스의 유전체는 조각나지 않은 하나의 가닥 형태의 RNA(non-segmented, single strand, positive-sense RNA)로 되어 있습니다. 전체 유전체의 크기는 대략 15kb(kilobases)이며, 유전자를 읽어서 단백질을 만들 수 있는 부위인 ‘ORF(open reading frame)’가 최소 10개로 구성되어 있습니다.

ORF 1a와 1b의 정보를 읽어 pp1a와 pp1b라는 두 개의 다단백질(poly-protein)을 만들어내고, 이후 바이러스의 14개 비구조 단백질(Nsp’s, non-structural proteins)로 쪼개집니다. 비구조 단백질은 바이러스의 형태를 구성하지 않지만 증식 과정에서 활용되는 단백질들을 말하며, PRRS 바이러스의 경우 4 종류의 단백질 분해 효소(proteases)와 RNA 중합효소(RNA-dependent RNA polymerase), 나선효소(helicase) 등이 비구조 단백질에 해당됩니다.
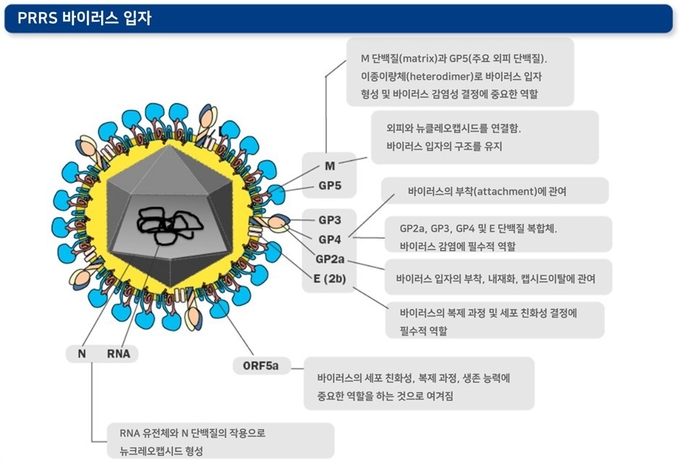
ORF 2-7은 8개의 구조 단백질(structural proteins)을 만들어냅니다(ORF2a, ORF2b, ORF3, ORF4, ORF5, ORF5a, ORF6, ORF7). 구조 단백질은 메이져 구조 단백질과 마이너 구조 단백질로 분류됩니다.
● 메이져 구조 단백질: N-글리코실화(N-당화) 되어있는 GP2a, GP3, GP4 등으로 구성된 삼량체 외피 단백질(trimeric envelop protein) 및 비(非)글리코실화 단백질인 2b(또는 E 단백질)이 해당됩니다. 삼량체 단백질의 경우 단독 작용 또는 GP5와의 복합 작용으로 바이러스의 감염성을 결정하는 중요한 역할을 합니다. E 단백질 역시 삼량체 단백질과 작용하여 바이러스의 증식 및 특이적인 세포 친화성을 결정합니다.
● 마이너 구조 단백질: 주요 외피 단백질인 GP5, 비글리코실화 막단백질 M, 뉴클레오캡시드 단백질 N 등이 해당됩니다. GP5와 M 단백질은 이종이량체(heterodimer)를 형성하여 바이러스 입자의 형성 및 바이러스의 감염에 필수적인 역할을 합니다. 또한, 5a라고 불리는 단백질은 바이러스의 세포 친화성, 증식 과정, 생존 능력에 중요한 역할을 하는 것으로 여겨집니다. ORF7이 만들어내는 N 단백질은 바이러스의 RNA와 상호작용하여 감염성 있는 바이러스 입자를 생성합니다. 이 부분은 진단 분석을 위한 항원으로 널리 이용되는데, 항원성이 크고 발현량이 많기 때문입니다.

PRRS 바이러스의 RNA는 유전체의 역할을 하면서 바이러스의 mRNA로도 작용합니다. 즉 PRRS 바이러스는 직접 단백질을 만들어낼 수 있습니다.
PRRS 바이러스의 구조
PRRS 바이러스는 45-80nm 직경의 구형의 입자입니다. 최근 초저온-전자 단층 촬영(cryo-electron tomography) 연구를 통해 바이러스의 뉴클레오캡시드가 기존에 예상했던 대칭 형태가 아니라 비대칭적인 형태일 것이라는 가설이 제기되고 있습니다. N 단백질과 RNA 유전체로 구성된 뉴클레오캡시드는 지질을 함유하는 외피로 둘러싸여 있습니다.
PRRS 바이러스의 물리화학적 특성은?
PRRS 바이러스는 RNA 바이러스이기 때문에, 상당히 불안정한 것으로 여겨집니다. PRRS 바이러스는 가온이나 건조 과정을 통해 쉽게 불활화될 수 있습니다. PRRS 통제 전략을 세울 때뿐 아니라 진단을 위한 가검물 운송 시에도 이러한 바이러스의 특징을 염두에 두는 것이 좋습니다.

PRRS 바이러스는 페놀, 포름알데히드, 대부분의 세제 및 소독제에 의해 불활화됩니다. 또한 pH 변화에 매우 민감하여 pH 6 이하 또는 7.5 이상에서 쉽게 감염성을 소실합니다.
그러므로, 농장에서 일상적으로 실시하는 수세 및 소독 절차는 환경 중의 PRRS 바이러스를 충분히 불활화할 수 있습니다. 요오드(0.0075%, 1분간), 4급 암모늄(0.0063%, 1분간), 염소(0.03%, 10분간) 계열 화학물을 이용하면 바이러스를 완전히 불활화할 수 있습니다. 농장 반입 물품 표면의 PRRS 바이러스 역시 10분간 자외선을 조사하면 완벽히 불활화할 수 있습니다.

참고 문헌
-
Allende R, Laegreid WW, Kutish GF, Galeota JA, Wills RW, Osorio FA. Porcine reproductive and respiratory syndrome virus: description of persistence in individual pigs upon experimental infection. J Virol. 2000, 74:10834-7.
-
Allende R, Kutish GF, Laegreid W, Lu Z, Lewis TL, Rock DL, Friesen J, Galeota JA, Doster AR, Osorio FA. Mutations in the genome of porcine reproductive and respiratory syndrome virus responsible for the attenuation phenotype. Arch Virol. 2000, 145:1149-61.
-
Brockmeier SL, Loving CL, Palmer MV, Spear A, Nicholson TL, Faaberg KS, Lager KM. Comparison of Asian porcine high fever disease isolates of porcine reproductive and respiratory syndrome virus to United States isolates for their ability to cause disease and secondary bacterial infection in swine. Vet Microbiol. 2017, 203:6-17.
-
Chang CC, Yoon KJ, Zimmerman JJ, Harmon KM, Dixon PM, Dvorak CM, Murtaugh MP. Evolution of porcine reproductive and respiratory syndrome virus during sequential passages in pigs. J Virol. 2002, 76:4750-63.
-
Cortey M, Díaz I, Martín-Valls GE, Mateu E. Next-generation sequencing as a tool for the study of Porcine reproductive and respiratory syndrome virus (PRRSV) macro- and micro- molecular epidemiology. Vet Microbiol. 2017. doi: 10.1016/j.vetmic.2017.02.002.
-
Darwich L, Gimeno M Sibila M, Diaz I, de la Torre E, Dotti S, Kuzemtseva L, Martin M, Pujols J, Mateu E. Genetic and immunobiological diversities of porcine reproductive and respiratory syndrome genotype I strains. Vet Microbiol. 2011, 150:49-62.
-
Dea S, Gagnon CA, Mardassi H, Pirzadeh B, Rogan D. Current knowledge on the structural proteins of porcine reproductive and respiratory syndrome (PRRS) virus: comparison of the North American and European isolates. Arch Virol. 2000, 145:659-88.
-
Díaz I, Gimeno M, Darwich L, Navarro N, Kuzemtseva L, López S, Galindo I, Segalés J, Martín M, Pujols J, Mateu E. Characterization of homologous and heterologous adaptive immune responses in porcine reproductive and respiratory syndrome virus infection. Vet Res. 2012, 19:43:30.
-
Dokland T. The structural biology of PRRSV. Virus Res. 2010, 154:86-97.
-
Domingo E, Holland JJ. RNA virus mutations and fitness for survival. Annu Rev Microbiol. 1997, 51:151-78
-
Dunowska M1, Biggs PJ, Zheng T, Perrott MR. Identification of a novel nidovirus associated with a neurological disease of the Australian brushtail possum (Trichosurus vulpecula). Vet Microbiol. 2012, 156:418-24.
-
Frydas IS, Nauwynck HJ. Replication characteristics of eight virulent and two attenuated genotype 1 and 2 porcine reproductive and respiratory syndrome virus (PRRSV) strains in nasal mucosa explants. Vet Microbiol. 2016, 182:156-62.
-
Frydas IS, Trus I, Kvisgaard LK, Bonckaert C, Reddy VR, Li Y, Larsen LE, Nauwynck HJ. Different clinical, virological, serological and tissue tropism outcomes of two new and one old Belgian type 1 subtype 1 porcine reproductive and respiratory virus (PRRSV) isolates. Vet Res. 2015, 46:37.
-
Gimeno M, Darwich L, Diaz I, de la Torre E, Pujols J, Martín M, Inumaru S, Cano E, Domingo M, Montoya M, Mateu E. Cytokine profiles and phenotype regulation of antigen presenting cells by genotype-I porcine reproductive and respiratory syndrome virus isolates. Vet Res. 2011, 18:42:9.
-
Goldberg TL1, Lowe JF, Milburn SM, Firkins LD. Quasispecies variation of porcine reproductive and respiratory syndrome virus during natural infection. Virology. 2003, 317:197-207.
-
Halbur P, Bush E. Update on abortion storms and sow mortality. Swine Health Prod. 1997, 5:73.
-
Han J, Wang Y, Faaberg KS. Complete genome analysis of RFLP 184 isolates of porcine reproductive and respiratory syndrome virus. Virus Res. 2006, 122:175-82.
-
Virus taxonomy: classification and nomenclature of viruses: Ninth Report of the International Committee on Taxonomy of Viruses. Ed: King AMQ, Adams MJ, Carstens EB, Lefkowitz EJ. Elsevier Academic Press. 2012.
-
Johnson CR, Griggs TF, Gnanandarajah J, Murtaugh MP. Novel structural protein in porcine reproductive and respiratory syndrome virus encoded by an alternative ORF5 present in all arteriviruses. J Gen Virol. 2011, 92:1107-16.
-
Le Gall A, Albina E, Magar R, Gauthier JP. Antigenic variability of porcine reproductive and respiratory syndrome (PRRS) virus isolates. Influence of virus passage in pig. Vet Res. 1997, 28:247-57.
-
Kapur V, Elam MR, Pawlovich TM, Murtaugh MP. Genetic variation in porcine reproductive and respiratory syndrome virus isolates in the midwestern United States. J Gen Virol. 1996, 77:1271-6.
-
Kuhn JH, Lauck M, Bailey AL, Shchetinin AM, Vishnevskaya TV, Bào Y, Ng TF, LeBreton M, Schneider BS, Gillis A, Tamoufe U, Diffo Jle D, Takuo JM, Kondov NO, Coffey LL, Wolfe ND, Delwart E, Clawson AN, Postnikova E, Bollinger L, Lackemeyer MG, Radoshitzky SR, Palacios G, Wada J, Shevtsova ZV, Jahrling PB, Lapin BA, Deriabin PG, Dunowska M, Alkhovsky SV, Rogers J, Friedrich TC, O’Connor DH, Goldberg TL. Reorganization and expansion of the nidoviral family Arteriviridae. Arch Virol. 2016, 161:755-68.
-
Lee C, Yoo D. The small envelope protein of porcine reproductive and respiratory syndrome virus possesses ion channel protein-like properties. Virology. 2006, 355:30-43.
-
Martelli P, Cordioli P, Fallacara F, Gozio S, Terreni M, Cavirani S. A follow up study of recurrent acute PRRS (Atypical PRRS-SAMS) and genetic variations of ORF5. In Proceedings of the 4th International Symposium on Emerging and Re-emerging Pig Diseases, 75-76, 2003, Italy.
-
Martín-Valls GE, Kvisgaard LK, Tello M, Darwich L, Cortey M, Burgara-Estrella AJ, Hernández J, Larsen LE, Mateu E. Analysis of ORF5 and full-length genome sequences of porcine reproductive and respiratory syndrome virus isolates of genotypes 1 and 2 retrieved worldwide provides evidence that recombination is a common phenomenon and may produce mosaic isolates. J Virol. 2014, 88:3170-81.
-
Mengeling WL, Lager KM, Vorwald AC. Clinical consequences of exposing pregnant gilts to strains of porcine reproductive and respiratory syndrome (PRRS) virus isolated from field cases of “atypical” PRRS. Am J Vet Res. 1998, 59:1540–4.
-
Meulenberg JJ, Petersen-den Besten A, De Kluyver EP, Moormann RJ, Schaaper WM, Wensvoort G. Characterization of proteins encoded by ORFs 2 to 7 of Lelystad virus. Virology. 1995, 206:155-63.
-
Meulenberg JJ, Petersen den Besten A, de Kluyver E, van Nieuwstadt A, Wensvoort G, Moormann RJ. Molecular characterization of Lelystad virus. Vet Microbiol. 1997, 55:197-202.
-
Morgan SB, Frossard JP, Pallares FJ, Gough J, Stadejek T, Graham SP, Steinbach F, Drew TW, Salguero FJ. Pathology and virus distribution in the lung and lymphoid tissues of pigs experimentally inoculated with three distinct type 1 prrs virus isolates of varying pathogenicity. Transbound Emerg Dis. 2014. doi: 10.1111/tbed.12272.
-
Morgan SB, Graham SP, Salguero FJ, Sánchez Cordón PJ, Mokhtar H, Rebel JM, Weesendorp E, Bodman-Smith KB, Steinbach F, Frossard JP. Increased pathogenicity of European porcine reproductive and respiratory syndrome virus is associated with enhanced adaptive responses and viral clearance. Vet Microbiol. 2013, 163: 13-22.
-
Murtaugh MP, Elam MR, Kakach LT. Comparison of the structural protein coding sequences of the VR-2332 and Lelystad virus strains of the PRRS virus. Arch Virol. 1995, 140:1451-60.
-
Murtaugh MP, Yuan S, Faaberg KS. Appearance of novel PRRSV isolates by recombination in the natural environment. Adv Exp Med Biol. 2001, 494:31-6.
-
Murtaugh MP, Stadejek T, Abrahante JE, Lam TT, Leung FC. The ever-expanding diversity of porcine reproductive and respiratory syndrome virus. Virus Res. 2010, 154:18-30.
-
Music N, Gagnon CA. The role of porcine reproductive and respiratory syndrome (PRRS) virus structural and non-structural proteins in virus pathogenesis. Anim Health Res Rev. 2010, 11:135-63.
-
Oleksiewicz MB, Stadejek T, Maćkiewicz Z, Porowski M, Pejsak Z. Discriminating between serological responses to European-genotype live vaccine and European-genotype field strains of porcine reproductive and respiratory syndrome virus (PRRSV) by peptide ELISA. J Virol Methods. 2005, 129:134-44.
-
Rowland RR, Steffen M, Ackerman T, Benfield DA. The evolution of porcine reproductive and respiratory syndrome virus: quasispecies and emergence of a virus subpopulation during infection of pigs with VR-2332. Virology. 1999, 259:262-6.
-
Shi M, Lam TT, Hon CC, Hui RK, Faaberg KS, Wennblom T, Murtaugh MP, Stadejek T, Leung FC. Molecular epidemiology of PRRSV: a phylogenetic perspective. Virus Res. 2010, 154:7-17.
-
Sinn LJ, Zieglowski L, Koinig H, Lamp B, Jansko B, Mößlacher G, Riedel C, Hennig-Pauka I, Rümenapf T. Characterization of two Austrian porcine reproductive and respiratory syndrome virus (PRRSV) field isolates reveals relationship to East Asian strains. Vet Res. 2016, 47:17.
-
Snijder EJ, Meulenberg JJ. The molecular biology of arteriviruses. J Gen Virol. 1998, 79:961-79. Snijder EJ, Dobbe JC, Spaan WJ. Heterodimerization of the two major proteins is essential for arterivirus infectivity. J Virol. 2003, 77:97-104. Snijder EJ, Kikkert M, Fang Y. Arterivirus molecular biology and pathogenesis. J Gen Virol. 2013, 94:2141-63.
-
Spilman MS, Welbon C, Nelson E, Dokland T. Cryo-electron tomography of porcine reproductive and respiratory syndrome virus: organization of the nucleocapsid. J Gen Virol. 2009, 90:527-35.
-
Stadejek T, Oleksiewicz MB, Potapchuk D, Podgórska K. Porcine reproductive and respiratory syndrome virus strains of exceptional diversity in Eastern Europe support the definition of new genetic subtypes. J Gen Virol. 2006, 87:1835-41.
-
Stadejek T, Stankevicius A, Murtaugh MP, Oleksiewicz MB. Molecular evolution of PRRSV in Europe: current state of play. Vet Microbiol. 2013, 165:21-8.
-
Sun L, Li Y, Liu R, Wang X, Gao F, Lin T, Huang T, Yao H, Tong G, Fan H, Wei Z, Yuan S. Porcine reproductive and respiratory syndrome virus ORF5a protein is essential for virus viability. Virus Res. 2013, 171:178-85.
-
Tian D, Wei Z, Zevenhoven-Dobbe JC, Liu R, Tong G, Snijder EJ, Yuan S. Arterivirus minor envelope proteins are a major determinant of viral tropism in cell culture. J Virol. 2012, 86:3701-12.
-
Truong HM, Lu Z, Kutish GF, Galeota J, Osorio FA, Pattnaik AK. A highly pathogenic porcine reproductive and respiratory syndrome virus generated from an infectious cDNA clone retains the in vivo virulence and transmissibility properties of the parental virus. Virology. 2004, 325:308–19.
-
Van Vugt JJ, Storgaard T, Oleksiewicz MB, Bøtner A. High frequency RNA recombination in porcine reproductive and respiratory syndrome virus occurs preferentially between parental sequences with high similarity. J Gen Virol. 2001, 82:2615-20.
-
Wang X, Marthaler D, Rovira A, Rossow S, Murtaugh MP. Emergence of a virulent porcine reproductive and respiratory syndrome virus in vaccinated herds in the United States. Virus Res. 2015, 210:34-41.
-
Weiland E, Wieczorek-Krohmer M, Kohl D, Conzelmann KK, Weiland F. Monoclonal antibodies to the GP5 of porcine reproductive and respiratory syndrome virus are more effective in virus neutralization than monoclonal antibodies to the GP4. Vet Microbiol. 1999, 66:171-86.
-
Wissink EH, Kroese MV, van Wijk HA, Rijsewijk FA, Meulenberg JJ, Rottier PJ. Envelope protein requirements for the assembly of infectious virions of porcine reproductive and respiratory syndrome virus. J Virol. 2005, 79:12495-506.
-
Zhao K, Ye C, Chang XB, Jiang CG, Wang SJ, Cai XH, Tong GZ, Tian ZJ, Shi M, An TQ. Importation and Recombination Are Responsible for the Latest Emergence of Highly Pathogenic porcine reproductive and respiratory syndrome virus in China. J Virol. 2015, 89:10712-6.
-
Zimmerman JJ, Benfield DA, Dee SA, Murtaugh MP, Stadejek T, Stevenson GW, Torremorell M. Porcine reproductive and respiratory syndrome virus (porcine arterivirus). In: 10th ed. Diseases of swine, Ed. Wiley-Blackwell. 2012, 31:463-86.
















